Gene expression patterns in a cell determine cellular specificity of development, differentiation and growth activity as well as cellular response or competency to virus, bacterial, and mycete. Whereas these expression patterns are regulated by many factors, we had been concentrating on following three factors; 1) transcriptional factors, which operate on the promoter region of their target gene, 2) chromatin remodeling factors that modulate the state of chromatin activity, 3) miRNA, which suppresses many genes at the post-transcriptional level. Joint Research Division of RNA Therapeutics with miRaX Therapeutics (Project for RNA Regulation) was established on July, 2020, and here, we will concentrate on elucidating how regulatory networks are disturbed in human diseases using strong inhibitors of specific miRNAs which we have developed.
Research interests
Development of RNA medicine using RNA decoys, TuD/S-TuD, which achieve strong and long-term suppression of specific microRNA.
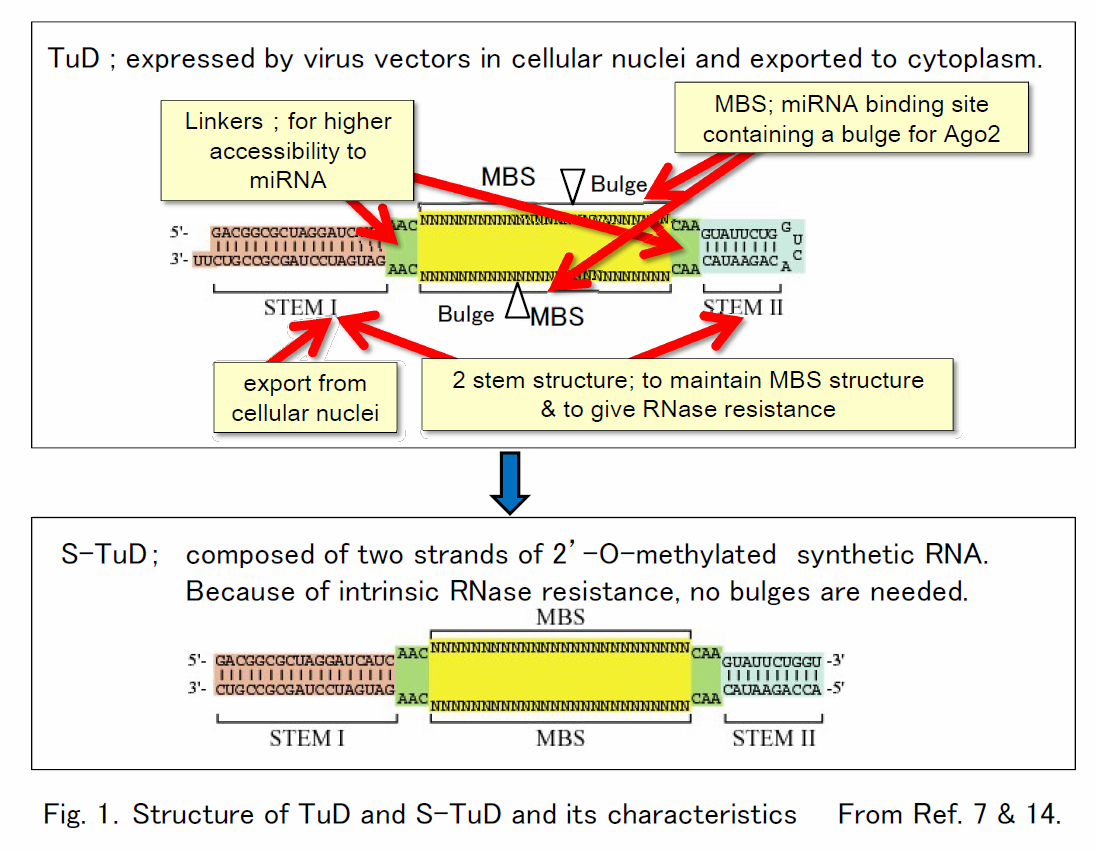
microRNA (miRNA) is a class of non-coding RNA, which suppresses a group of target mRNA by destabilizing them and/or their efficiency of translation. There are more than 2000 miRNAs in human. A single miRNA can suppress multiple mRNAs simultaneously, and it forms regulatory networks among many genes. Therefore each miRNA can function in development, differentiation, inflammation, immunity, or stress response and its abnormal expression could be causative of many human diseases including infectious disease and cancer. To understand these biological processes and to regulate these diseases, it is essential to have tools to strongly suppress specific miRNA for a long time. We have already established RNA decoy designated TuD (Tough Decoy), which suppress activity of specific miRNA when expressed by several virus vectors including those of retro/lentivirus and AAV. It has been confirmed by several third groups in Europe and USA that TuD has much higher inhibitory activity than other conventional methods.
Each TuD RNA molecule has a unique secondary structure containing 4 functional elements as shown in Fig 1. By mimicking the secondary structure of TuD, we also developed S-TuD (Synthetic TuD) composed of 2 strands of 2’-O-methyl synthetic RNA (Fig.1), which also exhibits strong inhibitory effects when introduced by transfection. We have applied TuD/S-TuD for the suppression of cancer stem cells in breast cancer, isolation and characterization of miRNAs involved in competency to HSV-1 infection (miR-199a-5p and -3p) and inhibition of passive cutaneous anaphylaxis (PCA) and allergic dermatitis using mouse models, and strong inhibitory effects of TuD/S-TuD are now being approved.
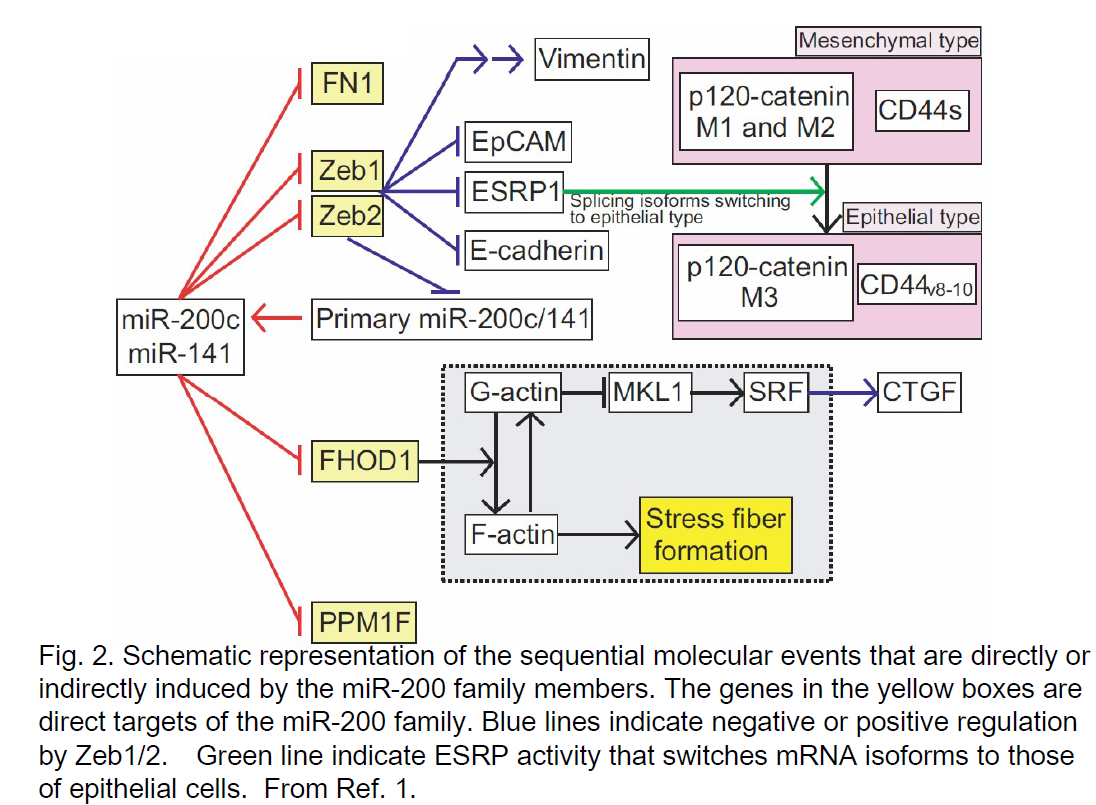
For the use of S-TuD as new therapeutic tools, elucidation of disease-specific gene regulatory networks and identification of most important target miRNA would be essential, and TuD/S-TuD are very useful even for such basic research. As for TuD, it is now improved by having two different MBS in a single TuD molecule. For example, by constructing such hybrid-type TuD, TuD-141/200c, we can now efficiently suppress the entire miR-200 family. By expressing TuD-141/200c by Tet-dependent pol III promoter, we show that epithelial-to-mesenchymal transition (EMT) of a colorectal tumor cell lines can be induced by continuous expression of the TuD for 2 weeks and, we can reverse the entire process and induce mesenchymal-to-epithelial transition (MET) by stopping TuD-141/200c expression. In Fig. 2, sequential molecular events that are propagated during this EMT were schematically summarized.
Kobayashi K*, Hiramatsu H*, Nakamura S, Kobayashi K, Haraguchi T, Iba H. (* equally contributed) Tumor suppression via inhibition of SWI/SNF complex-dependent NF-κB activation. Scientific Reports, 7:11772 (2017)
Kobayashi K, Suemasa F, Sagara H, Nakamura S, Ino Y, Kobayashi K, Hiramatsu H, Haraguchi T, Kurokawa K, Todo T, Nakano A, Iba H. MiR-199a inhibits secondary envelopment of Herpes Simplex Virus-1through the downregulation of Cdc42-specific GTPase activating protein localized in Golgi apparatus. Scientific Reports, 7:6650 (2017)
Hiramatsu H, Kobayashi K, Kobayashi K, Haraguchi T, Ino Y, Todo T, Iba H. The role of the SWI/SNF chromatin remodeling complex in maintaining the stemness of glioma initiating cells. Scientific Reports, 7:889 (2017)
Haraguchi T, Kondo M, Uchikawa R, Kobayashi K, Hiramatsu H, Kobayashi K, Chit UW, Shimizu T, Iba H. Dynamics and plasticity of the epithelial to mesenchymal transition induced by miR-200 family inhibition. Scientific Reports. 6:21117. (2016)
Kobayashi K, Sakurai K, Hiramatsu H, Inada K, Shiogama K, Nakamura S, Suemasa F, Kobayashi K, Imoto S, Haraguchi T, Ito H, Ishizaka A, Tsutsumi Y, Iba H. The miR-199a/Brm/EGR1 axis is a determinant of anchorage-independent growth in epithelial tumor cell lines. Scientific Reports, 5:8428 (2015)
Mizutani T, Ishizaka A, Suzuki Y, Iba H. 7SK small nuclear ribonucleoprotein complex is recruited to the HIV-1 promoter via short viral transcripts. FEBS Letters, 588:1630-1636 (2014)
Siddesha J.M, Valente A.J, Yoshida T, Sakamuri S.S.V.P, Delafontaine P, Iba H, Noda M, Chandrasekar B. Docosahexaenoic acid reverses angiotensin II-induced RECK suppression and cardiac fibroblast migration. Cellular Signalling, 26:933-941 (2014)
Kurashima Y, Amiya T, Nochi T, Fujisawa K, Haraguchi T, Iba H, Tsutsui H, Sato S, Nakajima S, Iijima H, Kubo M, Kunisawa J, Kiyono H. Extracellular ATP mediates mast cell-dependent intestinal inflammation through P2X7 purinoceptors. Nature Communications, 3:1034 (2012)
Ishizaka A, Mizutani T, Kobayashi K, Tando T, Sakurai K, Fujiwara T, Iba H. Double PHD finger proteins DPF3a and 3b are required as transcriptional coactivators in the SWI/SNF complex-dependent activation of the NF-kB RelA/p50 heterodimer. J. Biol. Chem. 287:11924-11933 (2012)
Haraguchi T, Nakano H, Tagawa T, Ohki T, Ueno Y, Yoshida T, Iba H. A potent 2’-O-methylated RNA-based microRNA inhibitor with unique secondary structures. Nucleic Acids Res. 40:e58 (2012)
Hikichi M, Kidokoro M, Haraguchi T, Iba H, Shida H, Tahara H, Nakamura T. MicroRNA regulation of glycoprotein B5R in oncolytic vaccinia virus reduces viral pathogenicity without impairing its antitumor efficacy. Molecular Therapy, 19: 1107-1115 (2011)
Sakurai K, Furukawa C, Haraguchi T, Inada K, Shiogama K, Tagawa T, Fujita S, Ueno Y, Ogata A, Ito M, Tsutsumi Y, Iba H. microRNAs miR-199a-5p and -3p target the Brm subunit of SWI/SNF to generate a double-negative feedback loop in a variety of human cancers. Cancer Res. 71:1680-1689 (2010)
Tando T, Ishizaka A, Watanabe H, Ito T, Iida S, Haraguchi T, Mizutani T, Izumi T, Isobe T, Akiyama T, Inoue J, Iba H. Requiem protein links RelB/p52 and the Brm-type SWI/SNF complex in a non-canonical NFκB pathway. J. Biol. Chem., 285: 21951-21960 (2010)
Mizutani T, Ishizaka A, Tomizawa M, Okazaki T, Yamamichi N, Kawana-Tachikawa A, Iwamoto A. Iba H. Loss of the Brm-type SWI/SNF chromatin remodeling complex is a strong barrier to the Tat-independent transcriptional elongation of HIV-1 transcripts. J. Virol., 83:11569-11580. (2009)
Yamamichi N, Shimomura R, Inada K, Sakurai K, Haraguchi T, Ozaki Y, Fujita S, Mizutani T, Furukawa C, Fujishiro M, Ichinose M, Shiogama K, Tsutsumi Y, Omata M, Iba H. LNA in situ hybridization analysis of miR-21 expression during colorectal cancer development. Clin. Cancer Res., 15:4009-4016. (2009)
Yamamichi N, Inada K, Furukawa C, Sakurai K, Tando T, Ishizaka A, Haraguchi T, Mizutani T, Fujishiro M, Shimomura R, Oka M, Ichinose M, Tsutsumi Y, Omata M, Iba H. Cdx2 and the Brm-type SWI/SNF complex cooperatively regulate villin expression in gastrointestinal cells. Exp. Cell Res., 315:1779-1789. (2009)
Haraguchi T, Ozaki Y, Iba H. Vectors expressing efficient RNA decoys achieve the long-term suppression of specific microRNA activity in mammalian cells. Nucleic Acids Res, 37:e43. (2009)
Fujita S, Ito T, Mizutani T, Minoguchi S, Yamamichi N, Sakurai K, Iba H. Mir-21 gene expression triggered by AP-1 is sustained through a double negative feedback mechanism. J. Mol. Biol., 378:492-504. (2008)
Minoguchi S, Iba H. Instability of retroviral DNA methylation in embryonic stem cells. Stem Cells, 26:1166-1173. (2008)
Kayama H, Ramirez-Carrozzi V. R, Yamamoto M, Mizutani T, Kuwata H, Iba H, Matsumoto M, Honda K, Smale S.T, Takeda K. Class-specific regulation of pro-inflammatory genes by MYD88 pathways and IκBζ. J. Biol. Chem., 283:12468-12477. (2008)
Fujita S, Iba H. Putative promoter regions of miRNA genes involved in evolutionarily conserved regulatory systems among vertebrates. Bioinformatics, 24:303-308. (2008)